
生命编码背后的故事(卅六)
中心法则

沃森回美国后参加的第一个学术活动是加州理工学院在1953年9月举办的国际会议。布拉格、佩鲁茨和肯德鲁以及欧洲和美国各地的蛋白质专家均应邀前来,在南加州这个小校园里济济一堂。只有牛津大学的霍奇金不得不缺席。因为思想激进左倾,她被美国外交部拒发签证。会议的主题是蛋白质,但与会者明白真正的目的在于庆祝鲍林的学术生涯。他们当然也都乐于躬逢其盛。克里克也从纽约赶来与老朋友相会。半年前,还在英国的沃森伙同彼特和多诺霍等人促狭地伪造一封鲍林致克里克极为热情的邀请信。克里克和布拉格都上了当,分别白白激动和沮丧一场。鲍林非常生气,以克扣每月给彼特的零用钱作为处罚。
蛋白质α螺旋结构中的遗留问题已经圆满解决。桑格对胰岛素中氨基酸序列的测定终于证实霍夫迈斯特和费舍尔半个世纪前提出的肽链一级结构。鲍林的α螺旋和β折叠二级结构也不再是空中楼阁。鲍林那年刚过52岁,无疑可以志得意满。但他私下心有戚戚。几个月前,他还打算着在会议上展示那时正在完善中的三螺旋DNA模型,让自己硕果累累的学术成就再上一层楼。然时过境迁,这个蛋白质会议上最引人注目的竟然是沃森随身带来的DNA双螺旋模型。
鲍林更大的烦恼还在象牙塔之外。他与艾娃计划在那年年底环球旅行,直到行前也没能拿到护照而不得不取消。他已经退出所有的政治性组织不再参与社会活动,还是只能以临时获取的限制性护照短暂出境。鲍林只好继续保持缄默,将环球旅行计划延后一年。
然而,1954年注定不平静。3月1日,美国在太平洋岛屿成功爆炸第一颗实用型氢弹,展示前所未有的威力。国内的麦卡锡主义甚嚣尘上。鲍林一度的老朋友、美国原子弹功臣奥本海默遭受审查,被剥夺接触国防机密的资格。面对日益严重的威胁,鲍林终于打破沉默,再度走向前台频频发表针砭时弊的演讲。他旗帜鲜明地反对继续制造、实验核武器,谴责政府对科学家和知识分子的迫害。10月时,他的护照申请又一次被拒绝。那时的鲍林处之泰然,不再去申诉。
几星期后,形势又有了变化。鲍林在康奈尔大学访问时得到喜讯,获得该年的诺贝尔化学奖。那是他等待良久的荣誉,尤其让他倍感欣慰的是诺贝尔奖委员会给出的获奖原因:“因为他对化学键本质的研究及其在阐明复杂物质结构中的应用”1。这个笼统的描述有违诺贝尔奖只奖励具体的“发现或发明”的传统和要求,更像是个“终身成就奖”。而“化学键的本质”确实是鲍林几十年心血结晶,远比那已经在DNA分子结构面前黯然失色的蛋白质结构“发现”更具科学意义。
鲍林的获奖众望所归,引起轰动的却是他的护照问题。瑞典的媒体公开猜测鲍林能否亲自来领奖。诺贝尔奖已经有半个世纪的历史,还只有纳粹德国政府曾经阻止公民领奖。被这么相提并论后,灰头土脸的美国外交部长不得不亲自干预,批准鲍林已经撤回的护照申请。这一次,他终于获得通用的普通护照,不再受时间和访问国别的限制。在斯德哥尔摩领奖后,鲍林和艾娃继续前往印度、日本等国,实现期望已久的环球旅行。

也是在那年,近30年前曾经在哥廷根为鲍林提供海森堡“不确定性原理”论文的量子力学创始人之一、72岁高龄的玻恩也终于等到自己姗姗来迟的诺贝尔物理奖。
克里克毕业离去后,布拉格也没能在卡文迪许实验室享受期盼已久的清净日子。他在1954年元旦辞职,踏着父亲30年前足迹去主持伦敦的王家研究所。伴随时代的进步,王家研究所历史悠久的圣诞节科学讲座已经成为电视实况转播的节目,影响今非昔比。63岁的布拉格与父亲当年一样乐此不疲。
在卡文迪许实验室接替布拉格的是物理学家莫特(Nevill Mott)。他的父母都曾是老主任汤姆森的部下,当年在这个实验室里相遇相爱。与布拉格不同,莫特对生物学进展漠不关心。当克里克为这位新主任介绍来访的沃森时,莫特惊诧莫名:“沃森?我一直以为你就是‘沃森-克里克’。”
莫特认为历史悠久的卡文迪许实验室应该延续汤姆森和卢瑟福奠定的传统,返回物理研究的正统。英国医学研究委员会那时也意识到他们长期资助的蛋白质研究项目栖身于这个物理实验室的尴尬。他们在剑桥投资兴建一个全新的分子生物学实验室,由佩鲁茨担任主任。他率领肯德鲁及研究血红蛋白的团队集体转移,同时也将桑格等剑桥生物化学人才纳入麾下。克里克也从美国回来加盟。他没有按计划去加州理工学院继续博士后研究。沃森一再警告洛杉矶的空气污染难以忍受。奥迪尔也怀上了他们的第二个女儿,希望回国分娩。拥有佩鲁茨、肯德鲁、桑格和克里克的新实验室阵容可观,立即成为正在脱颖而出的分子生物学众目所归的领头人。
卡文迪许实验室则在莫特领导下又一次改弦更张,致力于也是那个年代崛起的固体物理。莫特在20多年后赢得1977年的诺贝尔物理奖。他执掌的卡文迪许实验室雄风不减,只是早已与生命科学分道扬镳。
在美国的那一年里,克里克和沃森天各一方,只能通过频繁的信件继续他们旷日持久的争论。沃森在信中经常抱怨洛杉矶的天气和他研究RNA分子结构中遭遇的困难,也反复提起他那个“DNA --> RNA --> 蛋白质”的生命过程。他一直还没能说服好朋友接受这个发现DNA双螺旋结构之前的病中偶得。
在1954年2月13日的一封信中,沃森又画出更为具体的草图。他用箭头表示RNA可能来自DNA的化学变化;蛋白质则源自RNA,也许借助于类似伽莫夫想象的空穴机制出现。同时,DNA和RNA分子也可以通过它们的互补碱基对进行自我复制。这样,DNA中储存的编码信息可以逐步传送到RNA、蛋白质以及更多的DNA分子之中,主导生命的进程。沃森把他这个图像称之为“生命的奥秘”2。在加州理工学院,他已经说服物理学家费曼接受这个既疯狂又可爱的想法。连习惯质疑新思想的德尔布吕克也没有反对。
作为RNA领带俱乐部的“乐观者”,沃森名至实归。有着几个星期内成功破解DNA分子结构的经历和经验,他认定对付RNA无非探囊取物易如反掌。而RNA正是他那个生命奥秘中最关键的角色。但事与愿违,他和里奇全力以赴几个月,进展却乏善可陈。
同是米歇尔分离出的核酸,核糖核酸(RNA)与脱氧核糖核酸(DNA)的区别似乎只是其核糖分子中多出一颗未“脱落”的氧原子(见《十四:脱氧核糖》中的核糖分子结构图》),对核糖乃至整个核酸的结构影响不大。两种核酸中的碱基成分也略有不同。DNA有着腺嘌呤(A)、胞嘧啶(C)、鸟嘌呤(G)和胸腺嘧啶(T)。RNA中也有四种碱基,却是腺嘌呤(A)、胞嘧啶(C)、鸟嘌呤(G)和尿嘧啶(U)。二者之差只在于结构上相差不大的胸腺嘧啶和尿嘧啶(见《十一:分子结构》中的碱基分子结构图)。沃森因而认为RNA是由DNA经过简单的化学反应生成。但他们为RNA样品拍摄的X射线衍射照片总是十分模糊,无法与威尔金斯、富兰克林等人的DNA清晰照片相比。同时,RNA中的碱基成分比例也很不规范,有的符合与DNA相应的查戈夫定律(A = U;C = G),有的则完全没有规律。沃森不得不怀疑RNA分子没有特定的规则结构,更遑论简洁优美的双螺旋。
克里克也是RNA领带俱乐部当之无愧的“悲观者”。他早就预计到破解RNA分子结构的困难,对沃森和里奇的雄心壮志和研究方向不以为然。在菱形空穴猜想被证明不切实际后,伽莫夫和他的朋友泰勒还在继续寻找其它形状的空穴。克里克却已经认定任何在分子结构中为20种彼此相差无几的氨基酸分子量身定做几何模板的尝试都注定失败。他认为蛋白质的组装发生在带有编码的核酸——DNA或者RNA——分子之外,由另外的第三者帮忙实现。那会是一种小巧而特别的分子,像是连接电器元件时常见的“适配器”(adaptor)。它们一头可以连接特定的氨基酸分子。另一头则是编码该氨基酸分子的几个碱基分子。组装蛋白质时,也许DNA的双螺旋被打开,也许RNA只是一个单螺旋,它们之中的碱基分子都会顺着磷酸长链一字排开。一个个“适配器分子”在长链中找到与自己的碱基分子互补的位置,以氢键结合。在那同时,它们另一端所携带的氨基酸分子也就排列就绪,正好符合核酸分子中的编码次序。氨基酸分子彼此发生脱水缩合反应,遂成为崭新的蛋白质分子。这个机制简单直接,远比伽莫夫想象的空穴更为合理、灵活。
当然,适配器分子只存在于克里克的想象之中。从来没有人见识过如此巧夺天工的化学物。
伽莫夫的空穴要求DNA中的编码前后重叠关联,很快被克里克否证。克里克自己的适配器分子正好相反。它们彼此独立,单独地与核酸中一个或多个碱基分子互补连接。那正是氨基酸与碱基分子组合的“适配”。
碱基分子只有四个不同种类。如果适配器分子那一头只有一个碱基分子,这个机制只能用以区分四种不同的氨基酸。如果用两个碱基分子同时与核酸中碱基互补连接,它们会有16(4乘4)种不同组合,也不足以包容20种氨基酸。克里克因而认定适配器分子那一头肯定会有三个碱基分子,其不同组合多达64(4乘4乘4),代表20种氨基酸绰绰有余。也许它们并非一一对应,有着多个碱基组合编码同一种氨基酸。学物理出身的克里克从量子力学中借来现成名称,把这种重复性称为“简并”(degenerate)3。
这样一来,核酸中每三个邻近碱基分子合在一起代表一种氨基酸。沿着核酸的长链,一个个的碱基分子是三个一组的“莫尔斯电码”,每组对应一种氨基酸。通过适配器分子的运作,核酸中的碱基序列完全决定蛋白质中的氨基酸序列。那是两个平行等同——“共线性”(colinear)——的编码。
不同的氨基酸分子在蛋白质的肽链中会提供略为不同的氢键位置,让整个分子折叠出不同的形状。因此,中国人吴宪最早提出的蛋白质“折叠纯粹是一个氨基酸序列的功能。”4核酸中碱基分子和蛋白质中氨基酸分子的一维序列因而完全决定蛋白质的三维形状。而正如鲍林等人所揭示,蛋白质的外在形状是它们“一把钥匙开一把锁”式特异性功能的来源。千姿百态的蛋白质、丰富多彩的生物世界都源自各个特定的氨基酸序列和核酸中相应的碱基序列。
还在薛定谔叩问“生命是什么”之前,比德尔和塔特姆已经提出“一个基因一个酶”的猜测。那时候,基因还只是孟德尔和摩尔根分别从豌豆和果蝇遗传行为中总结的抽象概念。当沃森和克里克发现DNA的双螺旋结构后,他们立刻在论文中指出那中间以碱基对形式编码、可以自我复制的正是生命的基因信息。酶是蛋白质,所谓“一个基因一个酶”意味着每个蛋白质的氨基酸序列都来自DNA碱基序列中的相应基因信息。
1955年初,克里克在RNA领带俱乐部里分发题为《关于简并模板和适配器假说》5的论文,系统阐述他的想法。因为不是正式发表,论文语调轻松活泼,像是作者与朋友娓娓而谈的聊天记录。他一边指出伽莫夫模型的各种弊病,一边汲取其思想精华,脱胎换骨成为崭新的“适配器假说”。
伽莫夫没能为他的俱乐部募集到资助。成员们只好自己出钱领取人手一份的领带和领带夹,只在凑巧碰一起时聚个小会。在那之外,俱乐部不过是他们“群发”各自不成熟看法的松散“组织”。克里克如鱼得水,无所顾忌地接二连三推出一个又一个大胆的假设。在适配器假说一年后,他又与两位合作者一起有了重大发现。那两人之一是他在剑桥的老朋友,曾帮助他计算碱基分子氢键的格里菲斯。

他们考虑的是一个非常实际的问题。核酸中的编码可以用DNA中代表碱基的A、C、G、T或者RNA中相对应的A、C、G、U字母来表示。每三个碱基的组合——比如“ACG”“CGU”——代表一种氨基酸。于是,核酸的碱基长链是一个冗长的字母串,中间没有间隔。那如同中国古代的文言文,极难句读、理解。比如长链中有着“ACGACGACG”的序列时究竟是三个连续的“ACG”氨基酸还是其中有着两个相连的“CGA”或“GAC”氨基酸?那样稍有差错就会组合出完全不同的蛋白质,后果非同小可。莫尔斯设计电码时特意在长短电脉冲的组合之间加上适当的停顿,区分电文中每个字母的起始和终止。天然的核酸却不存在这样的方便。
克里克觉得自然的生命也应该有避免混淆的途径,比如“ACG”“CGA”“GAC”这三种组合中只能有一个是“有意义”(sense)的蛋白质编码。一旦确立后,其余两个均属于“无意义”(nonsense)。同样,三个字母完全重复的“AAA”“CCC”“GGG”“UUU”也因为会造成混淆而“无意义”。这样算计下来,四个字母以三个一组方式能构成的有意义组合不多不少,恰恰是20个。它们正好可以与20种氨基酸一一对应,不再有冗余的简并。
克里克很得意。在他眼里,这是一个浑然天成的“无逗号编码”6机制。
沃森花了很大的功夫才说服克里克接受他那个“DNA --> RNA --> 蛋白质”基因信息传送途径。那时,他已经改变注意,不再认为RNA来自DNA的化学反应。1956年,终于被说服的克里克又在俱乐部里发文,给出他自己绘制的版本。与沃森相同,克里克的图中的DNA和RNA可以分别自我复制,一代代地拷贝储存的信息。DNA的双螺旋打开时不仅可以复制如假包换的新DNA,也能够同样地构造出新的RNA。唯一的区别只是在后者中将胸腺嘧啶(T)置换为尿嘧啶(U)。DNA中的基因信息因而原封不动地复制给RNA,只是编码中的“T”被改为“U”。那是一个简单直接的信息“转录”(transcription)过程。
在细胞核外的细胞质中,DNA和RNA都有可能用自己的碱基编码组装蛋白质。这时,克里克大胆假设的适配器分子利用自己独特的结构将核酸中碱基序列“翻译”(translation)成氨基酸序列。组装后的蛋白质然后依照序列中的氢键位置自行折叠出各种各样的实用形状,成为生命运作中各司其职不可或缺的“元素”。
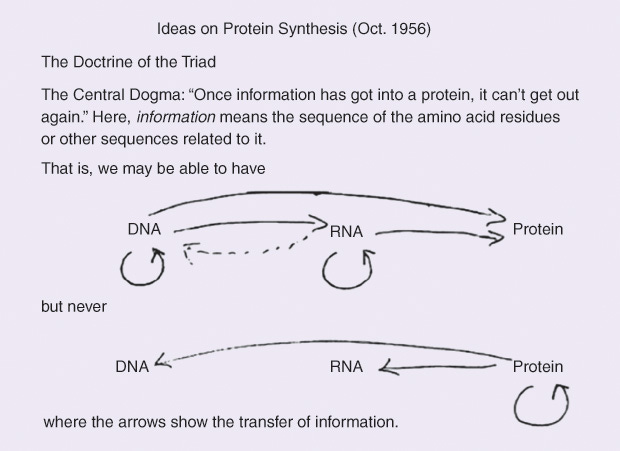
这个图像与沃森的想法大同小异。但克里克接下来着重强调这个信息传递过程的方向性。虽然他在自己的图中用虚线表示RNA中的基因信息有可能回馈给DNA,克里克指出蛋白质中的氨基酸序列绝不可能再将自己的信息传回DNA或RNA中的碱基序列。蛋白质也不能自我复制。它们必须由直接来自DNA或RNA中的信息组合而成。为此,他斩钉截铁地断言,以碱基或氨基酸序列编码的“信息一旦进入蛋白质,就不能再出来。”7蛋白质是基因信息的终极产物,犹如那时的物理学家尚未认识的“黑洞”(black hole)。
为突出这个论断的重要性,克里克特意赋予它一个掷地有声的名称:分子生物学的“中心法则”(central dogma)。储存有基因信息的DNA就像运筹帷幄的大元帅,将其指令传送RNA、制造各显神通的蛋白质。它却超然物外始终如一,完全不被自己启动、操纵的生命进程所左右。
这林林总总便构成沃森所谓的“生命奥秘”。
物理学家朗道以母牛和女孩为例讥讽李森科的故事在苏联不胫而走时,那里的知识界人士可能不会知道德国的魏斯曼早在19世纪末已经实际地做过朗道想象的实验。在李森科一手遮天的苏联,西方基因遗传学的知识已经荡然无存。魏斯曼曾一代代地割除小白鼠的尾巴,看到新生的小白鼠仍然长有尾巴。不过他只连续割过五代小白鼠尾巴,远不如人类处女膜的实例更具说服力,或更令人印象深刻。
魏斯曼的目的是验证达尔文的泛生假说。从古希腊的毕达哥拉斯到现代的达尔文,他们都曾设想生物体内各处会释放某种物质或信息,将毕生的经验教训凝聚于精子或卵子中传递给后代。那是李森科奉行的拉马克获得性遗传的生物学基础。魏斯曼的实验结果与朗道的直觉一致:小白鼠、母牛和女孩的遗传都没有受到后天经历的影响。魏斯曼由此提出生物的遗传完全取决于体内只有一套染色体的生殖细胞,即精子或卵子。它们不会受其它有着全数两套染色体的体细胞影响。
半个世纪后,克里克更进一步地指出:生物体内所有细胞——包括体细胞——的细胞核中的DNA都会保持原样,不因后天的生活经历改变。如果我们有办法直接读懂小白鼠DNA中的基因信息,就会看到它应该长有尾巴。即使那只小白鼠的尾巴在出生时已经被割除而未留痕迹。同样,一头拼命伸长脖子吃高处的树叶的鹿可能生长出更多的颈部肌肉和骨骼(蛋白质),却无力改变自己的DNA。后天的经历不仅无法遗传,也不能改变自己与生俱来的生命编码。
1957年9月19日,克里克在他母校伦敦大学学院的一次学术会议上发表演讲,首次将他一直还只是在RNA领带俱乐部中私下交流好几年的一系列新观点公诸于众。他只有一小时的发言时间,却需要涵盖太多的内容。即使是会场中的行家里手也无法跟上他口若悬河的语速。直到一年后他将演讲稿整理成论文发表后,这一整体思想才被人熟知。“中心法则”也随之成为分子生物学的一座里程碑。
然而在那个时刻,无论是中心法则、三个碱基(字母)组合的氨基酸编码、碱基和氨基酸序列的转录、翻译及共线性,还是适配器分子等等都还只是毫无事实根据的大胆假设。在1950年代渐入尾声之际,欧美各地的生物化学实验室纷纷投入小心求证的行列。
(待续)
for his research into the nature of the chemical bond and its application to the elucidation of the structure of complex substances
the mysteries of life
量子力学中,不同的物理状态可以有同样的能量,叫做“简并态”。
克里克语:"the folding is simply a function of the order of the amino acids" in the protein.
On Degenerate Template and the Adaptor Hypothesis
comma-free code
Once information has got into a protein it can't get out again.
学习到了科学发展的细节、进取。