
生命编码背后的故事(卅二)
核酸结构

鲍林和科里在论文中骄傲地宣布他们的模型史无前例,乃是“第一个精确描述的核酸结构”1。他们不知道克里克和沃森一年前的尝试,当然那个胎死腹中的探索也的确算不上先例。但鲍林绝对没想到他们的精确描述居然会与那两位小青年的构想如此相像。二者均是由三条磷酸链挤在中间的紧密结构,缺乏容纳水分子的空间。碱基分子一个个孤悬在外,不具备鲍林自己曾经绘声绘色描述过的基于互补的自我复制功能,当然更谈不上满足查戈夫的碱基成分比例定律。那次在邮轮上插肩而过后,鲍林还不知道查戈夫定律的存在。
克里克和沃森如获至宝。他们拿着那份预印本在各个实验室奔走相告,让剑桥的教授们为美国权威的阴沟翻船着实开心一番。然后,两人又到老鹰酒吧畅饮良久。
鲍林给儿子的信是他在1952年最后一天发出的很多封信之一。其它的信件分别奔向世界各地的学术名流,通报他的重大突破。在给兰德尔的长信中,鲍林特意为无缘得见国王学院的X射线衍射照片而不得不依赖阿斯特伯里和贝尔差强人意的老照片取得这一成就深表遗憾,不露声色地报了一箭之仇。
在同样焦躁地等待鲍林的论文时,威尔金斯碰上更大的惊奇。高斯林有一天突然找到威尔金斯,把那张标注“51号”的照片交给他保存。面对威尔金斯不解的询问,高斯林漫不经心地答曰:这都属于你的了,可以随意处置。
威尔金斯看到照片大为震惊。但在注意到拍摄日期后,他的激动很快转为极度不满。如此清晰的照片已经问世近九个月,富兰克林和高斯林却一直秘而不宣。威尔金斯既觉得不可思议也无可奈何,只能继续一天天地为富兰克林的离去数着日子。
1953年1月28日,富兰克林在国王学院举行告别讲座,总结她与高斯林在这里一年多的工作。在将近两小时的演讲中,富兰克林基本上只涉及用帕特森函数解析A型DNA结构的努力,对B型DNA鲜有提及。坐席中的威尔金斯非常失望。但让他更为焦虑不安的是富兰克林告而不别,讲座后依然没有要搬迁的意向——除了曾经让高斯林移交那张51号照片。
两天后,沃森又来到伦敦。他已经习惯在与合作者进行细菌性生活实验后顺道走访国王学院,与威尔金斯共进晚餐。这天,他随身带有鲍林论文的预印本。短短几天里,那篇论文已经成为他在同事间进行化学基础知识和智力测验的问卷,逢人拿出来看对方会用多久察觉出其中的几个谬误。不过他这次到来时威尔金斯正忙着,没空理他。兴冲冲的沃森遂转身闯进富兰克林的实验室,把正在昏暗中专心地用放大镜查看照相底片的富兰克林吓了一大跳。
鲍林给儿子寄预印本时也给卡文迪许实验室的布拉格寄了一份,但他没有同样地照顾国王学院的兰德尔。富兰克林听说鲍林的突破后立即给科里写信索取预印本,一直也没有回音。半年多前,科里在鲍林无法出境的情况下自己拜访国王学院时曾得到富兰克林的接待,看到过她拍摄的照片。富兰克林看到沃森带来的预印本后急忙翻阅,看到鲍林和科里只是在援引阿斯特伯里和贝尔的陈旧信息时随笔提及“威尔金斯的国王学院”还有更好的照片,完全没有提到她。
沃森没有察觉富兰克林的情绪变化,但也看出她没有兴致陪他玩智力测验游戏。于是他自顾自地喋喋不休起鲍林模型中的问题。富兰克林对这位趾高气扬的小年轻没有好气,两人随即陷入越来越激烈的争执。终于,沃森见势不妙夺门而出,在正好前来找他的威尔金斯掩护下全身而退。
看着沃森惊魂未定的狼狈样,威尔金斯惺惺相惜地控诉自己在富兰克林面前也曾有过相似的遭遇。回到他自己的办公室后,威尔金斯随手给沃森看他到手没几天的那张51号照片。沃森一见之下也目瞪口呆心跳加速。他已经亲手做过X射线衍射实验,不再像一年多以前对“观鸟者的傅立叶变换”一知半解。沃森领悟到照片中的“X”不仅无可辩驳地证明DNA具备螺旋结构,通过测量那“X”中条纹的距离还能够获知螺旋中长链的数目。
威尔金斯却没有心思讨论科学。两人一起去餐馆喝酒吃饭后,沃森独自乘坐夜班火车回剑桥。在寒冷的车厢里,沃森回味着他刚看过的照片,凭记忆将图形画在纸上。他像一年多前的克里克一样埋头估算,认定DNA的确不是他们那时想象的三螺旋,应该是一个简单得多、只有两条磷酸长链组成的双螺旋(double helix)。作为一群物理学家中唯一的生物学家,沃森觉得双螺旋更为合理。因为生物界最常见的东西总是成双成对。即使是物理学家的克里克也应该能接受这个基本常识。
第二天是星期六,沃森的兴奋还没有消退。他一大早又冲进佩鲁茨的办公室,在黑板上又写又画地描述他新得到的信息和想法。坐在角落里的布拉格这次对他重提DNA倒不以为许,颇有兴致地加入讨论。沃森早已深思熟虑,一针见血地指出鲍林在论文正式发表后肯定会很快认识到自己的错误。那只会激励他加倍努力,找出DNA的正确结构。卡文迪许实验室绝对不能只顾取笑而自己无所作为。这番言论重重地击中布拉格的软肋。他已经在晶体结构和蛋白质结构上两度惨败于鲍林,的确不愿意看到再而三。威尔金斯和富兰克林无休无止的内斗也让他对国王学院失去信心。卡文迪许实验室的人都知道,一年前他们作为媾和奉献的那些原子模型器具一直在国王学院某个角落积攒尘土,无人问津。鲍林的入局已经彻底改变DNA研究的格局。那不再是卡文迪许实验室、国王学院乃至英国医学研究委员会能够内部协调、切分的蛋糕。考量大局之后,布拉格当场拍板准许沃森重启DNA结构研究。
沃森特意留了个心眼,只用第一人称提议而没有兼顾克里克。他深知布拉格对那位“自己人”更为敏感,绝不会同意克里克在节骨眼上搁置论文再次误入歧途。得到恩准后,沃森大喜过望。他立即奔赴楼下的车间传达布拉格和佩鲁茨的旨意,要求他们以最快速度制作建造DNA模型所需的原子、分子模具。回办公室后,他又向克里克汇报他所看到的51号照片和自己的思考。克里克保持物理学家的本份,没有接受沃森的生物界成双成对奇谈怪论。他坚持在有确切证据之前,他们还必须兼顾双螺旋和三螺旋的可能性。
一星期后的2月8日,威尔金斯又一次应邀来到剑桥享用奥迪尔烹制的佳肴。他们的家宴已经有所扩大。沃森的妹妹蓓蒂在游历欧洲后来到这里,一边当保姆谋生一边与剑桥的英俊学生谈恋爱。彼特也像沃森一样经常前来混饭。
这一次,威尔金斯刚坐下就遭受克里克连番盘问,试图探知那张照片中更详细的数据。威尔金斯只顾狼吞虎咽,不置可否。饭后,沃森又拿出鲍林的预印本,让一直还没有机会拜读的威尔金斯浏览。威尔金斯在众目睽睽下逐一指出论文中谬误,顺利通过考试。沃森随后故伎重演地告诫威尔金斯:来自鲍林的威胁并未消失反而更为迫切。他必须立即开始建造DNA分子模型,否则必定还会被觉醒后的鲍林抢先。威尔金斯依然支支吾吾,解释说他不愿意贸然行事。一旦富兰克林辞行,他必定全力以赴。克里克打蛇随棍上地追询,那你不会介意我们先开始吧?
威尔金斯如梦初醒:这又是一次鸿门宴。
虽然很不情愿,威尔金斯还是绅士般地表示不会介意。他接着提醒两位咄咄逼人的新老朋友,国王学院年底前为英国医学研究委员会提供过一份总结报告,其中有他们想要的数据。
那晚,威尔金斯并不知道沃森早有布拉格的首肯,其实在一星期前已经开始了DNA的模型构造。但他也意识到形势比人强,自己的态度其实无关紧要。当他终于孤孤单单地告辞回家时,于心不忍的沃森追出来表示歉意。威尔金斯不再有心思搭理。
第二场鸿门宴的半年前,富兰克林的帕特森函数分析终于开始有所收获。A型DNA的X射线衍射图案导致一个出乎意料的结论:DNA——至少水分不多的A型DNA——具备一种“C2”对称性,对应于晶体学中常见的“面心立方”(face-centered cubic)结构。很多金属的晶体属于这种结构。它由最简单的立方体原胞重复排列成周期性的晶格,每个原胞的八个顶点和六个面中心都有一颗原子或分子。
如此简单明了的晶格结构显然与难以捉摸的螺旋不搭界。在一次少有的好心情中,富兰克林促挟地制作一幅四周带黑框的讣告,正式宣告A型DNA螺旋结构的寿终正寝。她和高斯林在上面签名后送给威尔金斯作为纪念。威尔金斯只觉得莫名其妙,克里克也嗤之以鼻。他们都笃信着DNA的螺旋结构。富兰克林其实也把握不定。从X射线衍射照片看来,A型和B型DNA迥然相异,压根看不出是同一个化合物。它们的实际区别却只不过是环境的湿度。如果DNA晶体在吸收、释放水分时从一个结构转变为另一个结构,那是物理上称作为“相变”(phase transition)的过程。DNA样品没有表现出任何相变迹象,不可能是在面心立方和螺旋这两个有着天壤之别的结构之间来回转换。因此,富兰克林一直没有正式发表这个结论,只在1952年底向医学研究委员会提交的报告中简要汇报。
为了促进被资助机构之间的交流,医学研究委员会内部设有一个具体协调、督查生物物理研究的领导小组。兰德尔和佩鲁茨都是小组成员。当克里克和沃森来探询国王学院的年终报告时,佩鲁茨不假思索地从文件堆中找出来给了他们。那不是公开发表的论文,但也没有保密要求。
国王学院的报告简明扼要,没有任何插图或照片。富兰克林的部分只占区区十来个段落。但克里克一看到DNA的C2对称性即豁然开朗。在研究蛋白质的卷曲螺旋时,他刚刚见识过这个对称性。具备这一对称性的不仅是面心立方晶体,也还有一种比较特别的螺旋结构。那正是沃森鼓吹的双螺旋,其中两条长链彼此不交错,表面上相互平行。但它们之中原子分子的排列顺序正好相反,因而是反平行。
尽管DNA在低湿度和高湿度环境下有着截然不同的X射线衍射图案,克里克认为二者其实是同样的螺旋结构。那既不是三股头发编结的辫子,也不是互相缠绕的树藤。它其实就是一座简单的“旋转楼梯”。楼梯两边的扶手便是两条反对称的长链,它们顺着楼梯走势盘旋而上——或一条“向上”一条“向下”。两条长链上以相反的次序交错着磷酸和脱氧核糖分子,形成反对称。在缺水的A型中,螺旋略像被压紧的弹簧,分子距离比较紧密而接近面心立方结构。吸收水分后,“弹簧”松开,不再如同面心立方而显露出以“X”形衍射图案为标志的螺旋真面目。国王学院的报告中还给出了具体的结构参数:低湿度的DNA吸水后沿着径向拉长,周期性由A型的2.8纳米变为B型的3.4纳米。
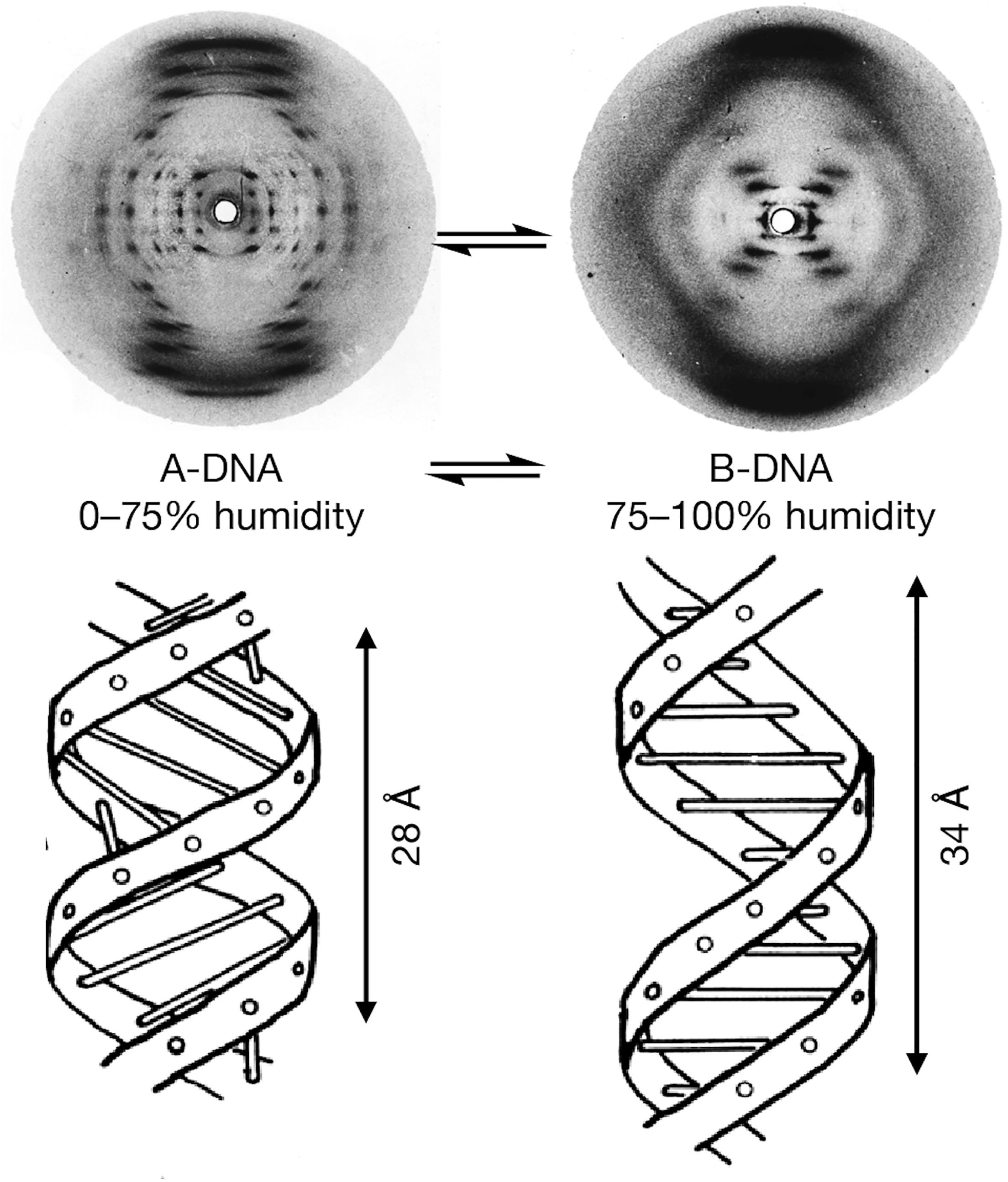
富兰克林早在一年多前的国王学院讲座中公开过这些数据,只是沃森那时既没有做笔记也压根没听明白。这一次,他们有了确切的数据,不再只凭想象和勇气进行大胆的假设。克里克也终于认可DNA的确是双螺旋。不过他根据的是实在的物理对称性,并非沃森所谓生物界自然的成双成对。
在卡文迪许实验室那间小小的办公室里,五位年轻人各自的小桌子靠墙放置,中间另有一张公用的小桌。沃森把它作为搭建DNA分子模型的工作台。他需要的模具还没有到齐,只好继续因陋就简,用硬纸板剪裁出各个分子的形状。在这个1953年的2月,他只能单枪匹马地鼓捣。克里克还必须埋头于自己的博士论文。但他显然无法集中注意力,时不时地抬头观看沃森的忙活,大声地批评、建议。
沃森既没能领会克里克的反平行概念,也没有接受富兰克林和威尔金斯一再强调的磷酸必须在分子外围才能从容地吸收、释放水分观点。与一年多前的三螺旋同样,他固执地把双螺旋的两条磷酸长链置放于分子中心位置,让一个个碱基分子孤悬在外。经历多次失败之后,沃森才听从克里克的劝告,尝试把长链放在分子外围作为“楼梯扶手”,让碱基分子在它们之间作为梯级。他俩都已经忘却半年前格里菲斯做过的氢键计算和与查戈夫的那场争执。沃森想象的还是克里克最初的念头:两个相同的碱基分子组成对,可以在互相分离时自我复制。为了这个“好主意”,他还特意在给德尔布吕克的信中洋洋得意了一下。
但他很快走进死胡同。鲍林的蛋白质α螺旋是一个单螺旋。虽然蛋白质中的各种氨基酸分子大小不一,它们两两相连形成的肽链依然构成标准的螺旋形状。当克里克、沃森和鲍林先后构造DNA的三螺旋时,他们不约而同地将磷酸长链置放于分子中心,让碱基挂在外围。这样,各种分子的不同大小也不至于影响到长链本身的螺旋形状。然而,当长链成为外围的楼梯扶手时,它们之间的每一块梯级都必须有着同样的长度才能保持两边的扶手之间的距离恒定。假如梯级有长有短,扶手就会在长的梯级处被挤开,在短梯级处被拉窄。那样歪七扭八的扶手不是标准的螺旋形,不可能衍射出那张51号照片的清晰“X”图案。
而碱基分子有两种。嘧啶是一个与苯环相似的碳环结构,嘌呤却有一大一小两个碳环。两个嘌呤分子连成的碱基对显然比两个嘧啶分子的碱基对长出很多,无法排在一起作为梯级。
即使是让不同的碱基分子通过氢键连接交叉配对,他也无法找到具备相同长度的组合。
办公室里的多诺霍没有加入过克里克和沃森的讨论。他对基因一窍不通,也不了解核酸的成分。但听到沃森在抱怨碱基分子时,多诺霍不由得来了兴致。跟随鲍林浸淫化学键多年后,他对各种分子的形状、尺寸了如指掌,在卡文迪许实验室正是研究碱基分子。他只看了一眼沃森的硬纸板就指出它们的形状不正确。
沃森大吃一惊,赶紧翻出他作为根据的几本参考书。多诺霍却不屑一顾地指出所有教科书都错了。
化学家早已熟知同分异构体的存在:相同化学成分的分子可以有不同的结构。碱基分子也有着两种不同的结构,分别名为“烯醇构型”(enol)和“酮构型”(keto)。它们之间的区别只是一颗氢原子处在分子外围的位置不同。长期以来,化学家认为碱基属于更为特殊的“互变异构体”(tautomer)。这样的分子结构上不稳定。那颗氢原子总是不老实地在两个位置来回跳跃,分子也就随着在烯醇构型和酮构型之间反复变迁。
多诺霍的研究推翻了这个既有成见。他发现碱基分子其实相当稳定,基本上会处于酮构型。而教科书里提供的却都只是它们的烯醇构型。
这一切在沃森听来犹如天方夜谭。多诺霍比沃森年长八年,也比他更早获得博士学位。在那间小办公室里,多诺霍相对沉默寡言,显得沉稳老练。斟酌之下,沃森决定听从多诺霍的意见,改换他硬纸板碱基分子中那颗氢原子的位置。
2月28日一大早,沃森来到还是空无一人的办公室。他在桌面上清理出一片空地,继续像玩七巧板似地摆弄那些代表不同碱基分子的硬纸板。由烯醇构型改为酮构型的分子外在形状变化并不大,嘌呤分子对与嘧啶分子对的长度依然悬殊。沃森不得不放弃那个相同分子组对的“好主意”,再次尝试不同碱基分子之间的配对。猛然间,他注意到因为那颗氢原子位置的迁移,碱基通过氢键配对的格局发生巨大变化。腺嘌呤(A)和胸腺嘧啶(T)连接起来的整体形状与鸟嘌呤(G)和胞嘧啶(C)组成的碱基对在外观上几乎完全一致。如果以这两个碱基对作为梯级,旋转楼梯的两个扶手之间就能处处保持同样的距离,完美无缺。
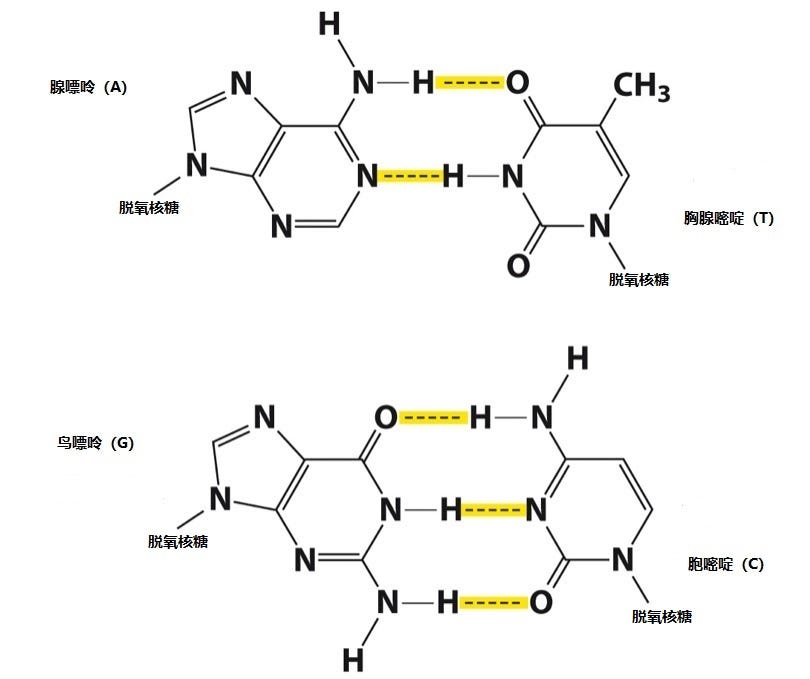
刚刚来上班的多诺霍在兴奋莫名的沃森注视下仔细检查那些硬纸板,确定没有问题。等到克里克终于现身时,沃森等不及他进屋就竹筒倒豆子般讲述这一重大进展。按照他俩的惯例,克里克在沃森激动情绪下保持着冷静的批评态度。他上上下下左左右右地审视那简陋的模型,却也没能挑出毛病。不仅如此,他看到A和T、G和C如此形成碱基对的两边都有现成的化学键与长链中的脱氧核糖相连。如果将这些碱基对翻转过来变成T和A、C和G组合,相应的化学键没有变化,完全可以同样地与长链相接。那正是他一直强调、沃森未能理解的反对称,可以满足富兰克林发现的C2对称性。克里克这才大松一口气,拍着沃森的后背表示祝贺。
这个新的模型里,两条由磷酸和脱氧核糖交错连接的长链构成DNA双螺旋的外围“扶手”。在它们之间,每个脱氧核糖分子连接着一个碱基分子,可以是腺嘌呤(A)、胞嘧啶(C)、鸟嘌呤(G)、胸腺嘧啶(T)中任何一个。但如果左边的碱基是A,与之相邻的右边必然会是T;如果左边是C,右边便是G。反之亦然:左边的T肯定伴随着右边的A,左边的G会带着右边的C。这样一来,腺嘌呤和胸腺嘧啶必定会同时出现,鸟嘌呤和胞嘧啶也同样地如影随形。那正是查戈夫早已发现的定律:A = T;C = G。
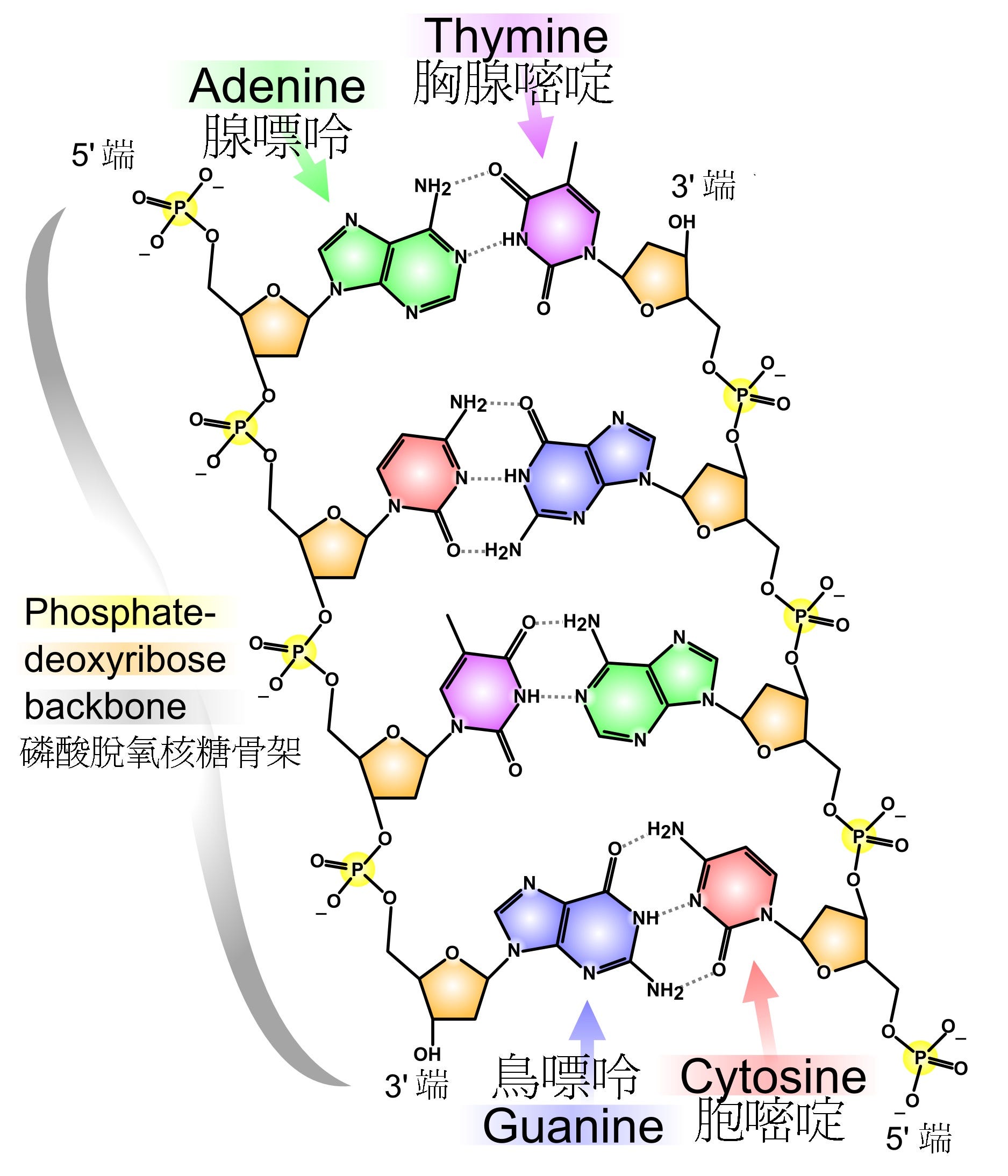
正如格里菲斯做过的计算,这些碱基分子互相以氢键连接,形成一个个的梯级拴住两边的长链扶手成为一个整体。磷酸长链彼此相距较远,不再需要带正电荷的金属离子搭桥。整个DNA分子宽松稀疏,有足够的空间容纳水分子。因为长度的限制和相应的查戈夫定律,作为梯级的碱基对只有四种可能的组合:A-T、C-G、G-C,T-A。无论以整体几何形状还是化学键位置而言,这四个碱基对几乎完全雷同,可以随意置换。它们因而能够在DNA长链中以任何顺序出现。虽然碱基对由不同的碱基分子组成,其中的两个碱基一一对应,正符合鲍林和德尔布吕克早年推测的以互补关系实现自我复制的功能。
如同莫尔斯电码中的长、短电脉冲,或现代计算机中的“0”和“1”,DNA长链中的一连串碱基对因而可以看作是由A、C、G、T四个“字母”——碱基分子对——编写出的文本。几乎毫无疑问,那应该就是生命发育、生长、遗传所需的指令信息。
整整十五年后,沃森在他题为《双螺旋》2的回忆录中生动地记叙那一历史时刻。在意识到这个DNA分子结构正是生命编码之所在后,两人即刻飞奔至老鹰酒吧。克里克当场向那里的午餐顾客宣告他们已经破解“生命的秘密”3。直至今日,老鹰酒吧门口设置着一块铭牌,纪念它作为发现生命秘密消息首发地点的荣耀。

这个激动人心的场景其实从来没有发生过。那只是沃森为追求戏剧性的杜撰。但在那个1953年2月28日的中午,照常在老鹰酒吧共进午餐的克里克和沃森的确切身感受着一个新时代的来临。
(待续)
the first precisely described strcuture for the nucleic acids that has been suggested by any investigator.
The Double Helix
the secret of life
花了 12 节总算讲了双螺旋,我认为太长,因为没有什么原理的进展,只是搞清楚细节而已;那些个人动作,最有用的是有关富兰克林对那三个人嗤之以鼻的态度;很多人替她抱怨说那些X光工作主要是她的,得奖却没份;但那个做模型工作她既然完全不相信甚至鄙视,工作做出了她没份很应该。